El MCA es una enfermedad genética rara de la retina causada por mutaciones genéticas en los genes OPN1LW, OPN1MW y LCR. Estos genes se encuentran en el cromosoma X y codifican proteínas llamadas fotopigmento, necesitado en los conos rojo y en los conos verde de la retina para captar la luz.
En esta página vemos cuáles son los genes responsables de la visión del color, cuáles las mutaciones genéticas que conducen a la enfermedad y la historia de dichos descubrimientos científicos en el campo de la genética molecular.
La BCM Families Foundation financia la investigación en genética molecular sobre el MCA y la creación de un registro de pacientes con MCA, para profundizar el conocimiento de las mutaciones genéticas que causan la enfermedad.
Los genes responsables de la visión del color
Las mutaciones genéticas en algunos genes que se hallan en el cromosoma X son las causantes del MCA. El cromosoma X se compone de dos brazos, el superior que se llama «p» y el inferior que se llama «q».
Los genes implicados en el MCA se encuentran en la localización Xq28, al final del brazo q.
En la imagen a continuación podemos ver el cromosoma X.
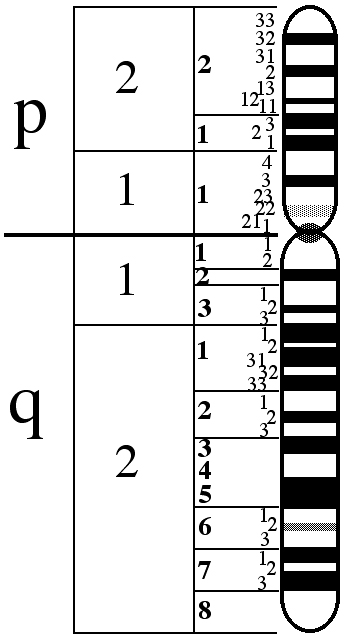
En la localización Xq28 se encuentran en el siguiente orden: los genes identificados con los nombres LCR, OPN1LW y OPN1MW.
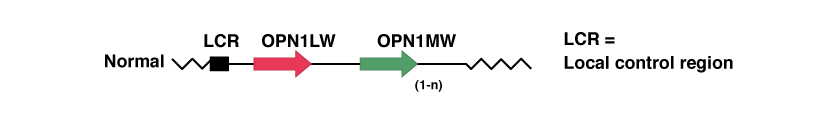
LCR es la región de control de locus (Locus Control Region en inglés), y actúa como promotora de la expresión de los dos genes siguientes. Ante la ausencia de este gen, ninguno de los dos siguientes se expresa en la retina humana. Además, la misma asegura que solo uno de los dos genes opsin (rojo y verde) se expresen de manera exclusiva en cada cono.
El OPN1LW y el OPN1MW son, respectivamente, los genes que contienen el código genético para las proteínas opsinas, es decir, las proteínas que constituyen los fotopigmentos para capturar la luz de color rojo (onda larga) y de color verde (onda media).
Muchas personas tienen varias réplicas del gen relacionado con el fotopigmento verde, el OPN1MW. Solo los dos primeros genes, pegados a la LCR, se expresan en la retina; las replicas sucesivas no se expresan. Por ejemplo, un varón caucásico tendrá en un 25 % de casos un gen OPN1MW único, en un 50 % tendrá 2 y en el restante 25 % de casos tendrá 3 o más.
En la imagen a continuación podemos ver una representación de estos genes en el caso de ausencia de mutaciones genéticas patológicas.
Para saber más sobre estos genes, se puede consultar la página de la National Center for Biotechnology Information (NCBI):
– OPN1LW, fotopigmento del cono rojo.
– OPN1MW, fotopigmento del cono verde.
– LCR, Locus Control Region, región de control de locus.
El gen responsable de la formación del fotopigmento azul se encuentra en una localización mucho más lejana: en el cromosoma 7, y el gen responsable de la formación de la rodopsina (el fotopigmento de los bastones) en el cromosoma 3:
– OPN1SW fotopigmento del cono azul.
– RODOPSINA fotopigmento de bastones.
En la siguiente imagen podemos ver las proteínas opsinas de los conos azul «S» (corto; short en inglés) y rojos «L» (largos).
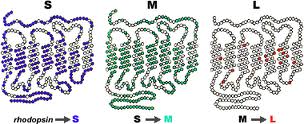
La imagen proviene de la página
Cogen la forma de una cadena que pasa siete veces dentro de un disco del segmento externo de un cono. Las tres proteínas son muy similares y, en particular, la M y la L se diferencian solo en algunos elementos que las componen. De hecho, ambos fotopigmentos, el rojo y el verde, son iguales en un 96 %, y sin embargo, se asemejan solo en un 46 % al fotopigmento azul.
Los genes OPN1LW y OPN1MW, como todos los genes, están formados por exones e intrones. En concreto, ambos genes tienen 6 exones, denominados de 1 a 6.
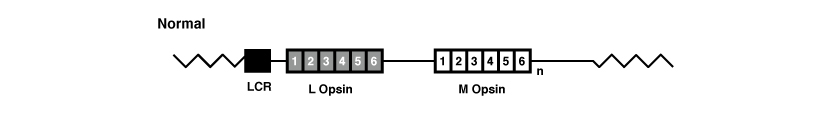
(La figura proviene de Jessica C. Gardner, Michel Michaelides, Graham E. Holder, Naheed Kanuga, Tom R. Webb, John D. Mollon, Anthony T. Moore, Alison J. Hardcastle ‘Blue cone monochromacy: Causative mutations and associated phenotypesMolecular Vision 2009; 15:876-884).
Como todas las proteínas, también las opsinas son estructuras tridimensionales que se doblan para tomar la forma tridimensional final. Algunos aminoácidos particulares presentes en la proteína son los responsables de que la misma se doble.
Mutaciones genéticas
Existen muchos tipos de mutaciones diferentes que pueden afectar al grupo de genes LCR, OPN1LW y OPN1MW.
Algunas mutaciones conducen a enfermedades llamadas comúnmente «daltonismo», cuyo defecto es solo la imposibilidad de diferenciar algunos colores.
Las mutaciones que conllevan el MCA identificadas hasta la fecha son las siguientes:
Grandes eliminaciones
1.
Eliminación de la LCR, o eliminación de la LCR y de algunos o todos los exones del gen OPN1LW.
Esta mutación es la ausencia de una gran parte de material genético. Al no estar presente el código genético de LCR, no se expresan las dos proteínas y, por lo tanto, los conos no poseen fotopigmentos rojos y verdes.
2.
Eliminación intragénica. Se trata de una eliminación de exones en el interior de los genes OPN1LW y OPN1MW, o la eliminación de material genético del primero y de parte del segundo.
Esta mutación también es la ausencia de una gran parte de material genético.
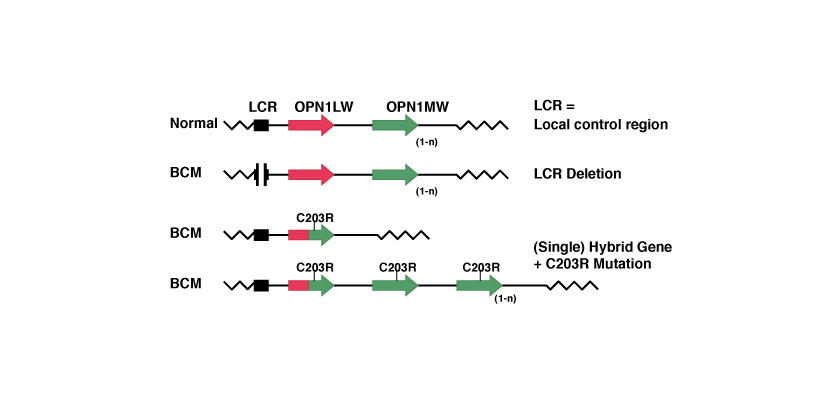
Mecanismo en dos fases de recombinación homóloga e inactivación.
En este caso, dada la similitud entre los dos genes OPN1LW y OPN1MW durante el proceso de «recombinación homóloga» se pierde uno de los dos genes y se crea un gen nuevo híbrido. Sucesivamente, una mutación puntual inactiva el gen restante.
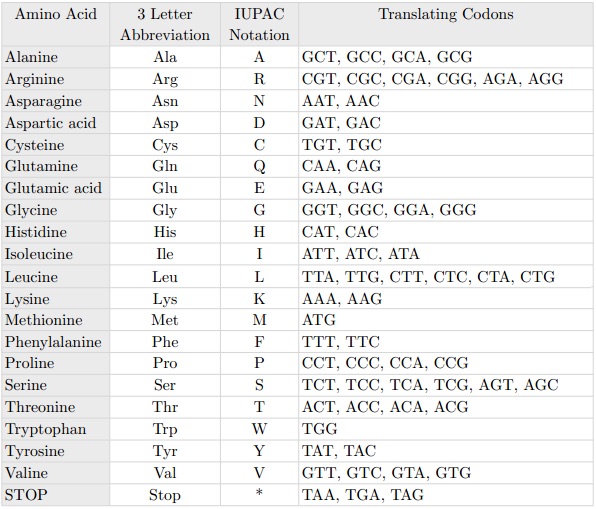
La mutación puntual más conocida es la llamada C203R. El nombre de la mutación puntual indica la posición donde se ha producido la mutación –en este caso, la localización 203– y qué aminoácido ha sido sustituido –en este caso la C de Cisteína con la R de Arginina. A nivel de los codones, se trata de una mutación puntual porque corresponde con la sustitución de la timina con la citosina en la localización 648, como podemos ver en la siguiente tabla:
La mutación C203R causa la proteina opsina una vez formada no se dobla en la forma tridimensional adecuada.
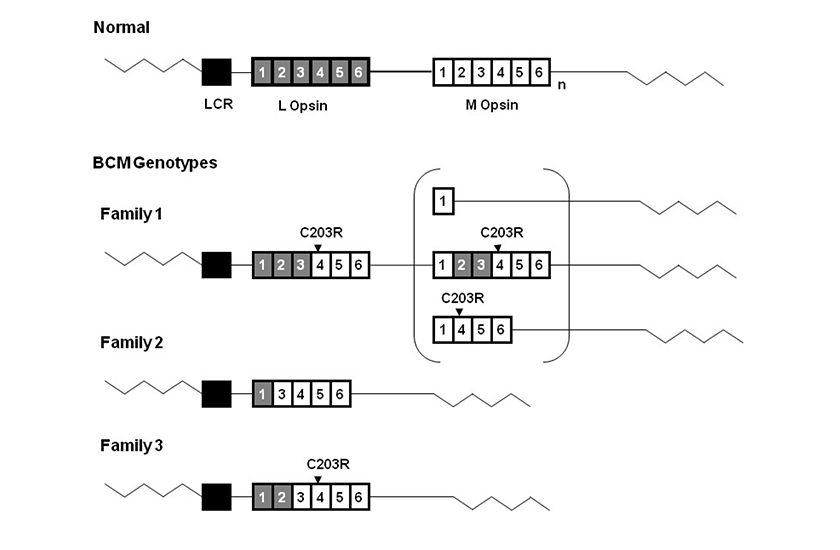
(La figura proviene de Jessica C. Gardner, Michel Michaelides, Graham E. Holder, Naheed Kanuga, Tom R. Webb, John D. Mollon, Anthony T. Moore, Alison J. Hardcastle ‘Blue cone monochromacy: Causative mutations and associated phenotypesMolecular Vision 2009; 15:876-884). Genotipos MCA.
Diagrama con fenotipos MCA de 3 familias británicas. La micromatriz L-M opsina tipo salvaje se muestra en la parte alta de la imagen. Los cuadrados grises representan los exones opsin L y los blancos representan los exones opsin M.
El subíndice «n» indica uno o más genes opsin M. Los cuadrados de color negro representan la región de control del locus (LCR). El LCR resulta presente sin la mutación en las tres familias. El punto de las mutaciones C203R detectado en la familia 1 y en la 3 se ve en la parte de arriba de los exones correspondientes. La familia 1 tiene un gen híbrido inactivo seguido por un segundo gen en la micromatriz. En el grupo se ven tres posibles estructuras de este segundo gen inactivo. La familia 2 tiene un gen híbrido no funcionante único, al cual le falta el exón 2. La familia 3 tiene un solo gen híbrido inactivo.
Otras mutaciones puntuales son la P307L, y la R247X. El ultimo reemplaza a arginina con un codón de terminación en la posición 247 que impide la formación de la proteína (mutación sinsentido).
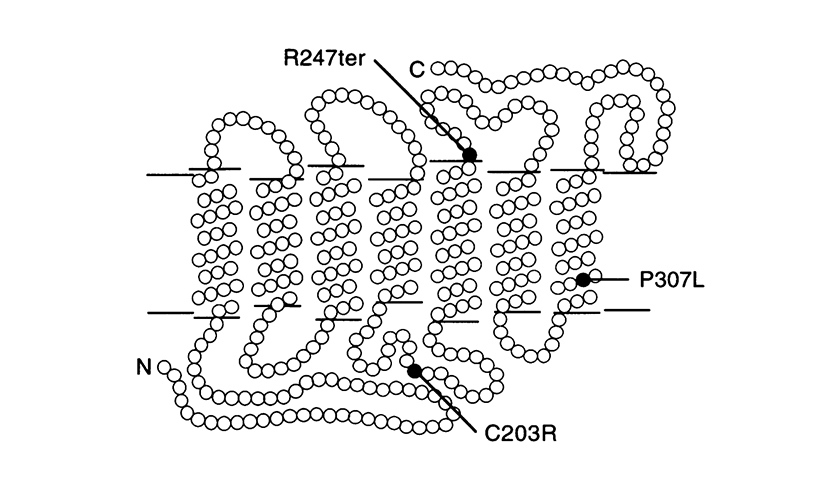
(La imagen proviene de Nathans et al. Am. J. Hum. Genet. 53: 987-1000, 1993.) Modelo de los pigmentos rojo y 5′ rojo – 2′ verde híbrido en la membrana fotoreceptora que muestra las localizaciones de las mutaciones puntuales identificadas en los monocromatos azules del cono. Cada punto representa un aminoácido. ‘N’ = amino terminal y ‘C’ = carboxilo terminal. El amino terminal está de cara al espacio extracelular.
Otras mutaciones
Otras mutaciones de los genes OPN1LW y OPN1MW que conducen al MCA representan un conjunto de mutaciones puntuales, llamadas, por ejemplo, LIAVA. Se padece el MCA si por culpa de una recombinación homóloga se produce, como en el caso anterior, un único gen mixto entre el OPN1LW y el OPN1MW y si el exón 3 contiene los siguientes aminoácidos en las localizaciones indicadas: Leucina 153, Isoleucina 171, Alanina 174, Valina 178 y Alanina 180. El nombre abreviado de este genotipo es LIAVA.
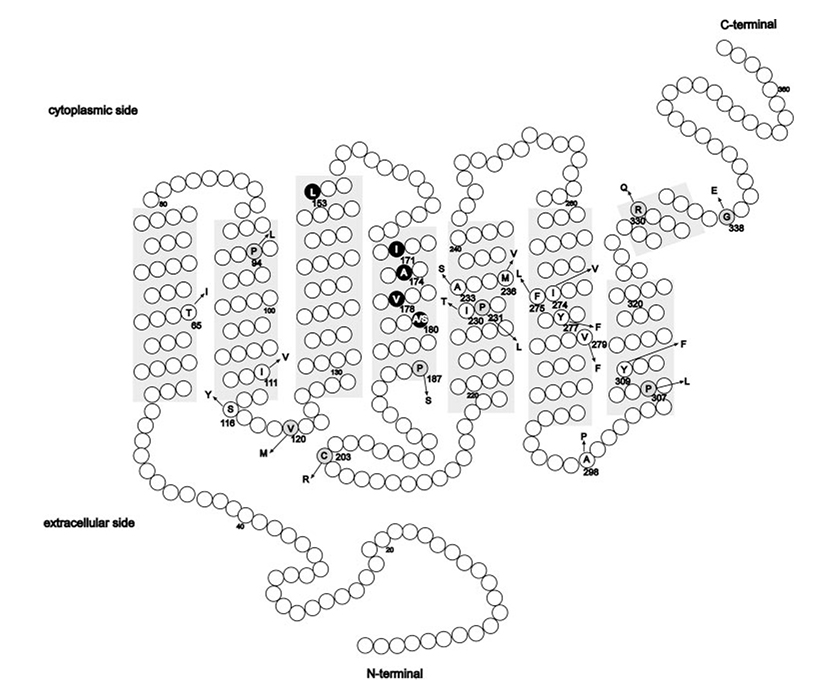
(La imagen proviene de Mizrahi-Meissonnier L., Merin S., Banin E., Sharon D., 2010). Las localizaciones de las alteraciones aminoácidas conocidas hasta la fecha se encuentran en los conos L y M de los genes opsin. Las zonas sombreadas representan los dominios de la transmembrana. Círculos: diferencias de los aminoácidos conocidas como polimorfismos con los aminoácidos más conocidos (con un código de una letra); flecha: variación del aminoácido. Podemos ver el número del codón para cada variación. Las variaciones sin sentido se asocian con una enfermedad relacionada con el cono opsin que puede causar el mal funcionamiento de la proteína y se ven representadas con el fondo gris. El haplotipo LIAVA aparece en negro.
| |
|
Nathans et al. 1993 |
| |
|
|
| |
|
Ayyagari et al. 2000 |
| |
A.Reitner, L.T. Sharpe, E. Zrenner 1991 |
|
| |
Mizrahi-Meissonnier L. et al. 2010 Neitz M. et al. 2004 Crognale MA. et al. 2004 |
Otras enfermedades con mutaciones genéticas en OPN1LW y OPN1MW
Otra enfermedad de la retina que se asocia a la localización Xq28 es la enfermedad de Bornholm, cuyos síntomas son similares a los del MCA. Se trata de una enfermedad muy rara y estacionaria. Para saber más, puede consultar la página web de OMIM y la página de la Universidad de Arizona.
Por último, cabe recordar que también existe una mutación particular de los dos genes OPN1LW y OPN1MW que causa una enfermedad diferente. Se trata de la mutación mal plegada, W177W, que, si está presente en ambos genes opsin, causa la distrofia de los conos con muestra de degeneración y apoptosis celular de los conos.
La historia del descubrimiento de los genes del MCA
Muchos investigadores han contribuido al descubrimiento de los genes involucrados en el MCA.
Cabe recordar los descubrimientos fundamentales de Jeremy Nathans sobre los genes responsables de la visión del color:
Nathans, J., Thomas, D., Hogness, D. S. Molecular genetics of human color vision: the genes encoding blue, green, and red pigments. Science 232: 193-202, 1986. PMID:2937147.
Nathans, J., Piantanida, T. P., Eddy, R. L., Shows, T. B., Hogness, D. S. Molecular genetics of inherited variation in human color vision. Science 232: 203-210, 1986. PMID: 3485310 .
Nathans, J. Molecular biology of visual pigments. Annu. Rev. Neurosci. 10: 163-194, 1987. PMID: 3551758 .
Nathans, J. The evolution and physiology of human color vision: insights from molecular genetic studies of visual pigments. Neuron. 24: 299-312, 1999. PMID: 10571225 .
Deeb, S. S. The molecular basis of variation in human color vision. Clin. Genet. 67: 369-377, 2005. PMID: 15811001 .
En particular, los trabajos que han permitido comprender la causa principal del monocromatismo de conos azules y, en concreto, el proceso en dos fases con mutación puntual de C2013R:
Nathans, J., Davenport, C. M., Maumenee, I. H., Lewis, R. A., Hejtmancik, J. F., Litt, M., Lovrien, E., Weleber, R., Bachynski, B., Zwas, F., Klingaman, R., Fishman, G. Molecular genetics of human blue cone monochromacy. Science 245: 831-838, 1989. PMID: 2788922.
Nathans, J., Maumenee, I. H., Zrenner, E., Sadowski, B., Sharpe, L. T., Lewis, R. A., Hansen, E., Rosenberg, T., Schwartz, M., Heckenlively, J. R., Traboulsi, E., Klingaman, R., Bech-Hansen, N. T., LaRoche, G. R., Pagon, R. A., Murphey, W. H., Weleber, R. G. Genetic heterogeneity among blue-cone monochromats. Am. J. Hum. Genet. 53: 987-1000, 1993. PMID: 8213841.
Reyniers, E., Van Thienen, M.-N., Meire, F., De Boulle, K., Devries, K., Kestelijn, P., Willems, P. J. Gene conversion between red and defective green opsin gene in blue cone monochromacy. Genomics 29: 323-328, 1995. PMID: 8666378.
Una investigación importante respecto a la mutación de tipo «eliminación» de la LCR o de la LCR y parte del gen OPN1LW:
Ayyagari, R., Kakuk, L. E., Bingham, E. L., Szczesny, J. J., Kemp, J., Toda, Y., Felius, J., Sieving, P. A. ‘Spectrum of color gene deletions and phenotype in patients with blue cone monochromacy’. Hum. Genet. 107: 75-82, 2000. Hum Genet. 2000 Jul;107(1):75-82. PMID: 10982039.
En lo que concierne a la eliminación intragénica recordamos el siguiente trabajo que identificó un caso de MCA con presencia del solo gen OPN1LW del fotopigmento rojo, pero con ausencia del exón 4 de este gen:
Ladekjaer-Mikkelsen, A.-S., Rosenberg, T., Jorgensen, A. L. ‘A new mechanism in blue cone monochromatism’. Hum. Genet. 98: 403-408, 1996. PMID: 8792812.
Reitner, A., Sharpe, L. T., Zrenner, E. Is colour vision possible with only rods and blue-sensitive cones? Nature 352: 798-800, 1991. PMID: 1881435 .
La Locus Control Region, y su papel en la expresión de genes opsin, fue el resultado de las siguientes obras:
Lewis, R. A., Holcomb, J. D., Bromley, W. C., Wilson, M. C., Roderick, T. H., Hejtmancik, J. F. Mapping X-linked ophthalmic diseases: III. Provisional assignment of the locus for blue cone monochromacy to Xq28. Arch. Ophthal. 105: 1055-1059, 1987. PMID: 2888453 .
Lewis, R. A., Nathans, J., Holcomb, J. D., Bromley, W. C., Roderick, T. H., Wilson, M. C., Hejtmancik, J. F. ‘Blue cone monochromacy: assignment of the locus to Xq28 and evidence for its molecular rearrangement’. Am. J. Hum. Genet. 41: A102 only, 1987.
Wang, Y., Macke, J. P., Merbs, S. L., Zack, D. J., Klaunberg, B., Bennett, J., Gearhart, J., Nathans, J. ‘A locus control region adjacent to the human red and green visual pigment genes’. Neuron 9: 429-440, 1992. PMID: 1524826.
En concreto, el papel que permite la expresión exclusiva de una opsina (roja o verde) en cada cono, ha sido descubierto en el último estudio.
Respecto al estudio de la mutación C203R, cabe recordar:
Kazmi MA, Sakmar TP, Ostrer H. ‘Mutation of a conserved cysteine in the X-linked cone opsins causes color vision deficiencies by disrupting protein folding and stablilty’. Investigative Ophthalmology and Visual Science. 1997; 38(6):1074–1081.
que agrupó los efectos negativos de esta mutación en el plegado de las proteínas opsinas y:
Winderickx J, Sanocki E, Lindsey DT, Teller DY, Motulsky AG, Deeb SS. Defective colour vision associated with a missense mutation in the human green visual pigment gene. Nature Genetics. 1992;1:251–256. PMID: 1302020
que estudió esta mutación y su frecuencia de alrededor del 2 % en las personas de origen caucásico.
En lo que concierne a las mutaciones raras tipo LIAVA se puede consultar:
Carroll J1, Neitz M, Hofer H, Neitz J, Williams DR., ‘Functional photoreceptor loss revealed with adaptive optics: an alternate cause of color blindness.’ Proc Natl Acad Sci U S A. 2004 Jun. PMID: 15148406.
Mizrahi-Meissonnier L., Merin S., Banin E., Sharon D., ‘Variable retinal phenotypes caused by mutations in the X-linked photopigment gene array’. Invest. Ophthalmol. Vis. Sci. 2010 Aug;51(8):3884-92. PMID: 20220053.
Neitz M, Carroll J, Renner A, et al. ‘Variety of genotypes in males diagnosed as dichromatic on a conventional clinical anomaloscope’. Vis Neurosci. 2004;21:205–216. PMID: 15518190.
Crognale MA, Fry M, Highsmith J, et al. ‘Characterization of a novel form of X-linked incomplete achromatopsia’. Vis Neurosci. 2004; 21:197–203. PMID: 15518189.
A continuación algunos de los primeros estudios de investigación sobre el MCA:
Huddart, J. ‘An account of persons who could not distinguish colours’. Phil. Trans. Roy. Soc. 67: 260 only, 1777.
Sloan, L. L. ‘Congenital achromatopsia: a report of 19 cases’. J. Ophthal. Soc. Am. 44: 117-128, 1954. PMID: 13131176.
Alpern, M., Falls, H. F., Lee, G. B. ‘The enigma of typical total monochromacy’. Am. J. Ophthal. 50: 996-1012, 1960. PMID: 13682677.
Blackwell, H. R., Blackwell, O. M. Rod and cone receptor mechanisms in typical and atypical congenital achromatopsia. Vision Res. 1: 62-107, 1961. https://doi.org/10.1016/0042-6989(61)90022-0.
Fleischman, J. A., O’Donnell, F. E. Jr. ‘Congenital X-linked incomplete achromatopsia. Evidence for slow progression, carrier fundus findings, and possible genetic linkage with glucose-6-phosphate dehydrogenase locus’. Arch Ophthalmol 1981;99:468-472. PMID: 6971088.
Lewis, R. A., Holcomb, J. D., Bromley, W. C., Wilson, M. C., Roderick, T. H., Hejtmancik, J. F. ‘Mapping X-linked ophthalmic diseases: III. Provisional assignment of the locus for blue cone monochromacy to Xq28′. Arch. Ophthal. 105: 1055-1059, 1987. PMID: 2888453.
En relación al estudio de la distrofia de conos, una enfermedad degenerativa debida a una mutuación puntual en ambos genes OPN1LW y OPN1MW:
Gardner JC, Webb TR, Kanuga N, Robson AG, Holder GE, Stockman A, Ripamonti C, Ebenezer ND, Ogun O, Devery S, Wright GA, Maher ER, Cheetham ME, Moore AT, Michaelides M and Hardcastle AJ,’X-Linked Cone Dystrophy Caused by Mutation of the Red and Green Cone Opsins’.The American Journal of Human Genetics 87, 26–39, July 9, 2010. PMID: 20579627.
Mencionamos también algunos estudios recapitulativos que tratan el tema:
Neitz J., Neitz M. ‘The genetics of normal and defective color vision’. 2011 Review. Vision Research. PMID: 21167193.
Deeb, S.S. ‘Molecular Genetics of colour vision deficiencies’. Clinical and Experimental Optometry 87.4 – 5 July 2004. PMID:15312026.