Blauzapfenmonochromasie ist eine seltene genetische Netzhauterkrankung, die durch genetische Mutationen in den Genen OPN1LW, OPN1MW und LCR hervorgerufen wird. Diese Gene befinden sich auf dem X-Chromosom und enthalten Proteine, so genannte Photopigment, die in den roten und grünen Zapfen benötigt werden, um Licht zu erfassen.
Hier sehen wir, welche Gene für die Farbwahrnehmung zuständig sind, welche genetischen Mutationen zur Erkrankung führen und welchen Entwicklungsweg diese molekulargenetischen Entdeckungen durchlaufen haben.
Die BCM Families Foundation unterstützt die molekulargenetische Forschung bei Blauzapfenmonochromasie und das Anlegen eines Blauzapfenmonochromaten-Registers, um die Erkenntnisse über die dieser Krankheit zugrunde liegenden genetischen Mutationen zu vertiefen.
Für die Farbwahrnehmung verantwortliche Gene
Blauzapfenmonochromasie wird durch genetische Mutationen an einigen Genen verursacht, die sich auf dem X-Chromosom befinden.
Das X-Chromosom besteht aus zwei Armen, wobei der kurze Arm mit p, der lange mit q bezeichnet wird.
Die an Blauzapfenmonochromasie beteiligten Gene befinden sich in der Position Xq28, am Ende des q-Arms.
In der nachstehenden Abbildung ist das X-Chromosom dargestellt.
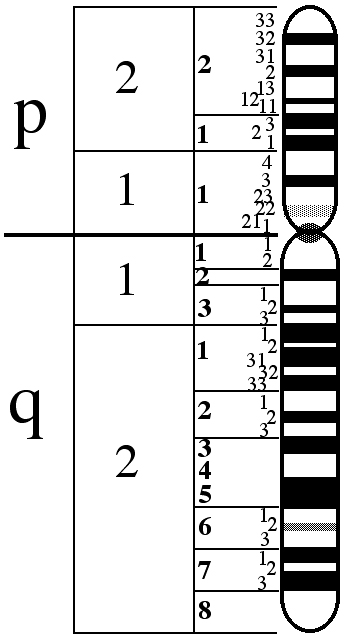
In der Position Xq28 befinden sich die Gene LCR, OPN1LW und OPN1MW in der genannten Reihenfolge.
LCR steht für Locus Control Region und fungiert als Expressionspromoter der beiden nachfolgenden Gene. Bei Fehlen dieses Gens wird keines der beiden nachfolgenden Gene in der menschlichen Netzhaut ausgedrückt. Darüber hinaus sorgt sie dafür, dass in den Zapfen jeweils ausschließlich eines der beiden Opsin-Gene (rot oder grün) ausgedrückt wird.
OPN1LW und OPN1MW sind Gene, die jeweils den genetischen Code für die OPSIN-Proteine enthalten, also die Proteine, die die Fotopigmente für die Absorption des roten (langwelligen) und grünen (mittelwelligen) Lichts bilden.
Viele Menschen haben mehrere Kopien des für das grüne Fotopigment zuständigen Gens OPN1MW. Aber nur die ersten beiden, unmittelbar hinter der LCR liegenden Gene werden in der Netzhaut ausgedrückt, während die nachfolgenden Kopien nicht ausgedrückt werden. So haben 25% der hellhäutigen Männer nur ein einziges OPN1MW-Gen, 50% haben 2 und die übrigen 25% haben 3 oder mehr.
In der nachfolgenden Abbildung sehen wir eine Darstellung dieser Gene, bei denen keine pathologischen genetischen Mutationen vorliegen.
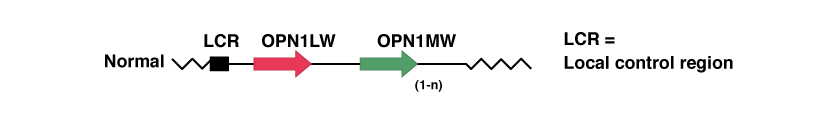
Zur Vertiefung der Kenntnisse über diese Gene empfiehlt sich die Website „National Center for Biotechnology Information“, NCBI:
– OPN1LW, Fotopigment der Rotzapfen;
– OPN1MW, Fotopigment der Grünzapfen;
– LCR, Locus Control Region.
Das Gen für die Bildung des blauen Fotopigments befindet sich in einer sehr weit entfernten Region auf dem Chromosom 7, während das für die Bildung von Rhodopsin (Fotopigment der Stäbchen) verantwortliche Gen auf dem Chromosom 3 angeordnet ist.
– OPN1SW Fotopigment der Blauzapfen;
– RHODOPSIN Fotopigment der Stäbchen.
In der nachfolgenden Abbildung sehen wir die OPSIN-Proteine der Blauzapfen des Typs „S“ (short), der Grünzapfen des Typs „M“ (medium) und der Rotzapfen des Typs „L“ (Long).
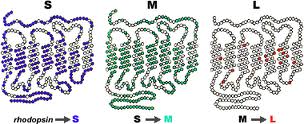
(Abbildung entnommen aus:
Sie haben die Form einer Kette, die 7-mal durch eine Scheibe des Außensegments eines Zapfens hindurchgeht. Die drei Proteine sind sich sehr ähnlich und insbesondere die Typen M und L unterscheiden sich nur in einigen Bestandteilen. Die roten und grünen Fotopigmente sind zu 96% identisch, während ihre Ähnlichkeit mit dem blauen Fotopigment nur bei 46% liegt.
Wie alle übrigen Gene bestehen auch OPN1LW und OPN1MW aus Exonen und Intronen. Beide Gene haben insbesondere 6 Exone, die von 1 bis 6 nummeriert sind.
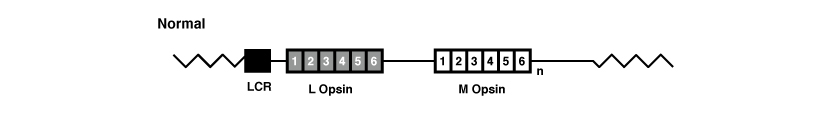
(Abbildung entnommen aus: Jessica C. Gardner, Michel Michaelides, Graham E. Holder, Naheed Kanuga, Tom R. Webb, John D. Mollon, Anthony T. Moore, Alison J. Hardcastle ‘Blue cone monochromacy: Causative mutations and associated phenotypes‘ Molecular Vision 2009; 15:876-884).
Wie alle übrigen Proteine sind auch OPSIN-Proteine dreidimensionale Gebilde, die gefaltet werden müssen, um ihre endgültige dreidimensionale Struktur anzunehmen. Einige besondere Aminosäuren dieses Proteins sind für dessen Faltung zuständig.
Genetische Mutationen
Es gibt zahlreiche genetische Mutationen, die die Gengruppen LCR, OPN1LW und OPN1MW betreffen können.
Einige Mutationen führen zu einer Krankheit, die gemeinhin als ‚Daltonismus‘ bezeichnet wird und deren einzige Auswirkung in der Unfähigkeit besteht, bestimmte Farben zu erkennen.
Bis jetzt wurden folgende Mutationen identifiziert, die Blauzapfenmonochromasie verursachen:
Große Deletionen
1.
LCR-Deletion bzw. Deletion der LCR und einiger oder aller Exone des Gens OPN1LW.
Bei dieser Mutation fehlt ein Großteil des genetischen Materials. Da der LCR-Code fehlt, werden die beiden Proteine nicht ausgedrückt und in den Zapfen sind folglich weder Rot- noch Grün-Fotopigmente enthalten.
2.
Intragene Deletion. Hierbei handelt es sich um eine Deletion der Exone im Innern der Gene OPN1LW und OPN1MW bzw. um eine Deletion des genetischen Materials des ersten und teilweise des zweiten Gens.
Auch bei dieser Mutation fehlt ein Großteil des genetischen Materials.
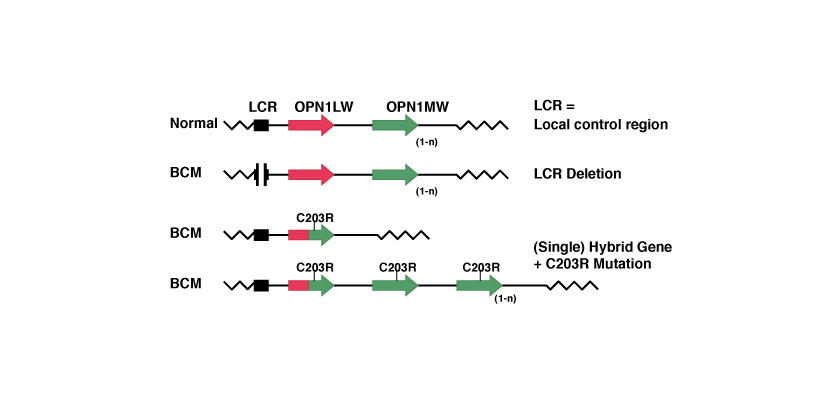
2-Step-Mechanismus mit homologer Neukombination und Inaktivierung
Aufgrund der zwischen den beiden Genen OPN1LW und OPN1MW bestehenden Ähnlichkeiten geht während der homologen Neukombination eines der beiden Gene verloren, während ein neues, hybrides Gen entsteht. Anschließend wird das verbliebene Gen durch eine Punktmutation inaktiviert.
Die bekannteste Punktmutation ist die sogenannte C203R-Mutation. Der Ausdruck Punktmutation weist auf die von der Mutation betroffene Position – hier 203 – sowie auf die ausgetauschte Aminosäure hin; in diesem Fall wurde C = Zystein durch R = Arginin ausgetauscht. Im Bereich der Codone ist dieser Austausch punktuell, da er einem Austausch von Thymin mit Zytosin an der Position 648 entspricht, wie in der folgenden Abbildung gezeigt wird:
Die C203R-Mutation bewirkt, dass sich das einmal gebildete Opsin-Protein nicht in die richtige dreidimensionale Form faltet.
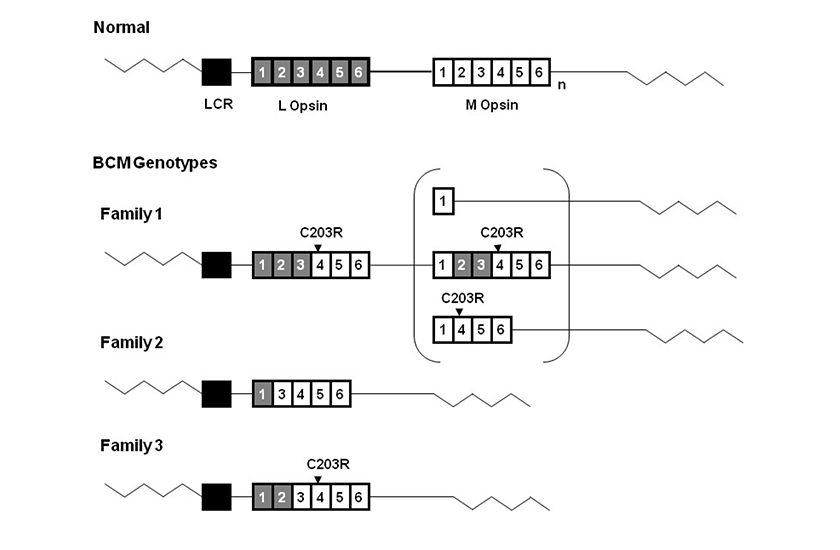
(Abbildung entnommen aus: Jessica C. Gardner, Michel Michaelides, Graham E. Holder, Naheed Kanuga, Tom R. Webb, John D. Mollon, Anthony T. Moore, Alison J. Hardcastle „Blue cone monochromacy: Causative mutations and associated phenotypes Molecular Vision“ 2009; 15:876-884). Blauzapfenmonochromasie-Genotypen. Diagramm zur Darstellung von Blauzapfenmonochromasie-Genotypen in 3 englischen Familien Der Wildtyp des L- und M-Opsin-Gen-Array ist in der Abbildung oben dargestellt. Graue Kästchen stellen L-Opsin-Exone dar, weiße Kästchen hingegen M-Opsin-Exone.
Ein tiefer gestelltes „n“ weist auf ein oder mehrere M-Opsin-Gene hin. Das schwarze Kästchen bezeichnet die Locus-Kontrollregion, LCR genannt. Die LCR war bei allen drei Familien ohne Mutation vorhanden. Die bei Familie 1 und Familie 2 festgestellten C203R-Punktmutationen sind über den entsprechenden Exonen dargestellt. Familie 1 hat ein inaktives, hybrides Gen, dem ein zweites Gen auf dem Array folgt. Die drei mögliche Strukturen dieses zweiten inaktiven Gens sind in der Klammer angegeben. Familie 2 hat ein einzelnes, nicht funktionsfähiges, hybrides Gen, bei dem Exon 2 fehlt. Familie 3 hat ein einzelnes inaktives hybrides Gen.
Weitere Punktmutationen sind P307L und R247X.
Letzteres ersetzt Arginin durch ein Stop-Codon an Position 247, das die Bildung des Proteins verhindert (Nonsense-Mutation).
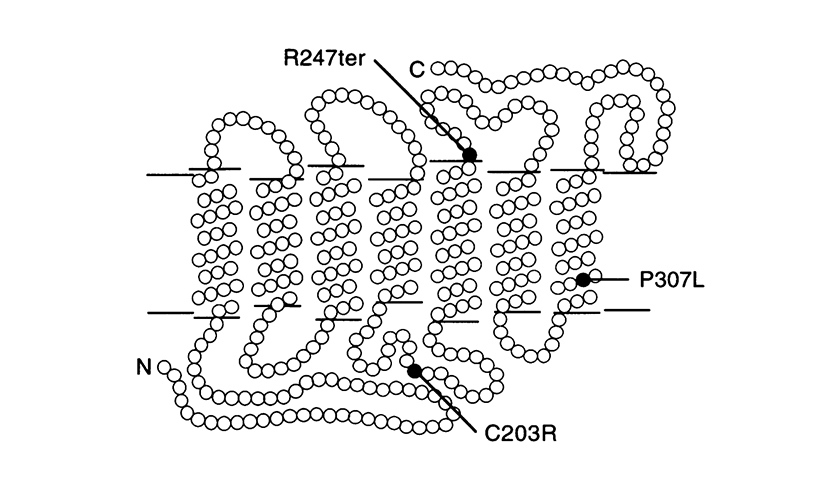
(Abbildung entnommen aus: J. Nathans et al. Am. J. Hum. Genet. 53: 987-1000, 1993.) Modell der hybriden Rot- und 5’-Rot – 2’-Grünpigmente in der Fotorezeptorenmembran mit Darstellung der Loci der bei Blauzapfenmonochromaten identifizierten Punktmutationen Jeder Kreis stellt eine Aminosäure dar. „N“ = Amino-Terminus und „C“ = Carboxy-Terminus Der Amino-Terminus steht vor dem extrazellulären Raum.
Weitere Mutationen
Weitere, zu Blauzapfenmonochromasie führende Mutationen in den Genen OPN1LW und OPN1MW bestehen aus einer Gruppe von Punktmutationen, die beispielsweise LIAVA genannt werden. Blauzapfenmonochromasie entsteht, wenn – wie im vorgenannten Fall – aufgrund einer homologen Neukombination ein einziges Misch-Gen aus OPN1LW und OPN1MW gebildet wird und das Exon 3 folgende Aminosäuren in den angegebenen Positionen enthält: Leuzin 153, Isoleuzin 171, Alanin 174, Valin 178 und Alanin 180. Dieser Genotyp wird mit dem Kürzel LIAVA bezeichnet.
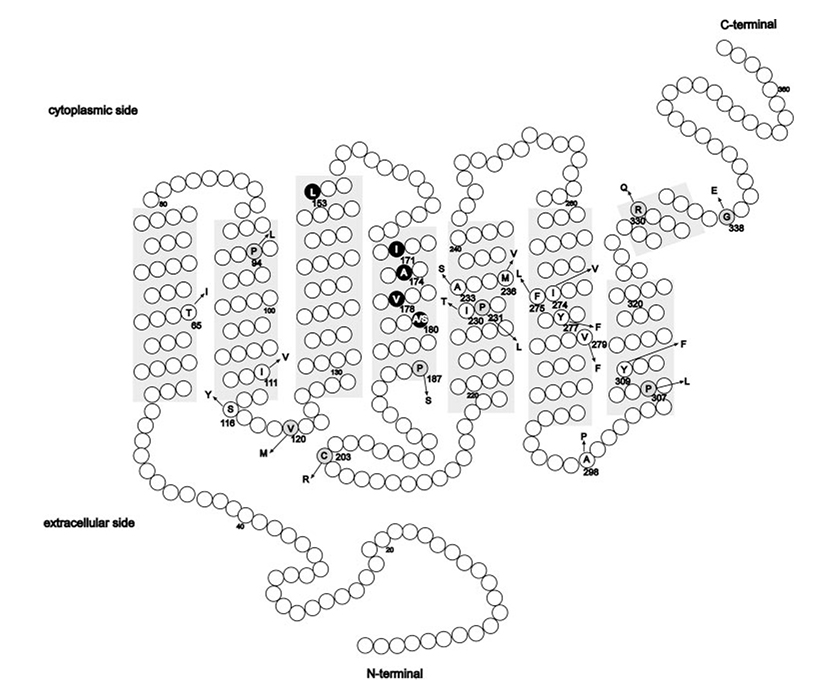
(Abbildung entnommen aus: Mizrahi-Meissonnier L., Merin S., Banin E., Sharon D., 2010.) Locus der Aminosäurenveränderungen in den Opsin-Genen der L- und M-Zapfen laut bisheriger Studien. Schattierte Bereiche: Transmembran-Domänen. Kreise: Aminosäurenunterschiede und bekannter Polymorphismus bei der gebäuchlichsten Aminosäure (in einbuchstabigem Code); Pfeil: Aminosäurenveränderung. Für jede Änderung ist die Codon-Nummer angegeben. Missense-Änderungen, verbunden mit einer durch Zapfen-Opsin verursachten Krankheit, die wahrscheinlich Proteindysfunktionen bewirken, sind grau unterlegt. Der LIAVA-Haplotyp ist schwarz hervorgehoben.
| |
|
Nathans et al. 1993 |
| |
|
|
| |
|
Ayyagari et al. 2000 |
| |
A.Reitner, L.T. Sharpe, E. Zrenner 1991 |
|
| |
Mizrahi-Meissonnier L. et al. 2010 Neitz M. et al. 2004 Crognale MA. et al. 2004 |
Weitere Krankheiten mit genetischen Mutationen in den Genen OPN1LW und OPN1MW
Eine weitere, mit der Position Xq28 verbundene Netzhauterkrankung ist die Bornholm-Augenkrankheit (BED), die ähnliche Symptome wie Blauzapfenmonochromasie aufweist. Es handelt sich um eine sehr seltene, stationäre Krankheit. Weiterführende Informationen finden sich unter OMIM sowie auf der Website der Universität Arizona.
Schließlich soll noch erwähnt werden, dass eine besondere Mutation in den beiden Genen OPN1LW und OPN1MW auch zu einer anderen Krankheit als Blauzapfenmonochromasie führen kann. Es handelt sich hierbei um die Fehlfaltungsmutation „W177R“. Wenn sie in beiden OPSIN-Genen auftritt, führt dies zu einer Zapfendystrophie mit erkennbarer Zelldegeneration und Zelltod.
Geschichte der Entdeckung der für Blauzapfenmonochromasie verantwortlichen Gene
Zahlreiche Forscher haben an der Entdeckung der an Blauzapfenmonochromasie beteiligten Gene mitgewirkt.
An dieser Stelle sei an die grundlegenden wissenschaftlichen Entdeckungen von Jeremy Nathans bei den für die Farbwahrnehmung verantwortlichen Genen erinnert:
Nathans, J., Thomas, D., Hogness, D. S. Molecular genetics of human color vision: the genes encoding blue, green, and red pigments. Science 232: 193-202, 1986. PMID:2937147.
Nathans, J., Piantanida, T. P., Eddy, R. L., Shows, T. B., Hogness, D. S. Molecular genetics of inherited variation in human color vision. Science 232: 203-210, 1986. PMID: 3485310 .
Nathans, J. Molecular biology of visual pigments. Annu. Rev. Neurosci. 10: 163-194, 1987. PMID: 3551758 .
Nathans, J. The evolution and physiology of human color vision: insights from molecular genetic studies of visual pigments. Neuron. 24: 299-312, 1999. PMID: 10571225 .
Deeb, S. S. The molecular basis of variation in human color vision. Clin. Genet. 67: 369-377, 2005. PMID: 15811001 .
Insbesondere an jene Arbeiten, die dazu beigetragen haben, die wichtigsten Ursachen der Blauzapfenmonochromasie und vor allem das 2-Step-Verfahren mit Punktmutation C203R aufzudecken:
Nathans, J., Davenport, C. M., Maumenee, I. H., Lewis, R. A., Hejtmancik, J. F., Litt, M., Lovrien, E., Weleber, R., Bachynski, B., Zwas, F., Klingaman, R., Fishman, G. Molecular genetics of human blue cone monochromacy. Science 245: 831-838, 1989. PMID: 2788922.
Nathans, J., Maumenee, I. H., Zrenner, E., Sadowski, B., Sharpe, L. T., Lewis, R. A., Hansen, E., Rosenberg, T., Schwartz, M., Heckenlively, J. R., Traboulsi, E., Klingaman, R., Bech-Hansen, N. T., LaRoche, G. R., Pagon, R. A., Murphey, W. H., Weleber, R. G. Genetic heterogeneity among blue-cone monochromats. Am. J. Hum. Genet. 53: 987-1000, 1993. PMID: 8213841.
Reyniers, E., Van Thienen, M.-N., Meire, F., De Boulle, K., Devries, K., Kestelijn, P., Willems, P. J. Gene conversion between red and defective green opsin gene in blue cone monochromacy. Genomics 29: 323-328, 1995. PMID: 8666378.
Eine weitere bedeutende Forschungsarbeit über Mutationen mit Deletion der LCR oder der LCR und eines Teils des Gens OPN1LW ist:
Ayyagari, R., Kakuk, L. E., Bingham, E. L., Szczesny, J. J., Kemp, J., Toda, Y., Felius, J., Sieving, P. A. ‘Spectrum of color gene deletions and phenotype in patients with blue cone monochromacy’. Hum. Genet. 107: 75-82, 2000. Hum Genet. 2000 Jul;107(1):75-82. PMID: 10982039.
Im Zusammenhang mit intragenen Deletionen erinnern wir an folgende Forschungsarbeit über einen Fall von Blauzapfenmonochromasie, bei dem nur das Gen OPN1LW des Rotpigments, aber ohne Exon 4 vorhanden war:
Ladekjaer-Mikkelsen, A.-S., Rosenberg, T., Jorgensen, A. L. ‘A new mechanism in blue cone monochromatism’. Hum. Genet. 98: 403-408, 1996. PMID: 8792812.
Reitner, A., Sharpe, L. T., Zrenner, E. Is colour vision possible with only rods and blue-sensitive cones? Nature 352: 798-800, 1991. PMID: 1881435 .
Der Locus-Kontrollregion, und seine Rolle in der Expression von Genen Opsin, war das Ergebnis der folgenden Arbeiten:
Lewis, R. A., Holcomb, J. D., Bromley, W. C., Wilson, M. C., Roderick, T. H., Hejtmancik, J. F. Mapping X-linked ophthalmic diseases: III. Provisional assignment of the locus for blue cone monochromacy to Xq28. Arch. Ophthal. 105: 1055-1059, 1987. PMID: 2888453 .
Lewis, R. A., Nathans, J., Holcomb, J. D., Bromley, W. C., Roderick, T. H., Wilson, M. C., Hejtmancik, J. F. ‘Blue cone monochromacy: assignment of the locus to Xq28 and evidence for its molecular rearrangement’. Am. J. Hum. Genet. 41: A102 only, 1987.
Wang, Y., Macke, J. P., Merbs, S. L., Zack, D. J., Klaunberg, B., Bennett, J., Gearhart, J., Nathans, J. ‘A locus control region adjacent to the human red and green visual pigment genes’. Neuron 9: 429-440, 1992. PMID: 1524826.
Der Grund dafür, dass einem Opsin (rot oder grün) in jedem Zapfen ausschließlich eine Expression möglich ist, wurde im letztgenannten Artikel entdeckt.
Im Zusammenhang mit der Erforschung der C203R-Mutation erinnern wir daran, dass in dieser Arbeit:
Kazmi MA, Sakmar TP, Ostrer H. ‚Mutation of a conserved cysteine in the X-linked cone opsins causes color vision deficiencies by disrupting protein folding and stablilty‘. Investigative Ophthalmology and Visual Science. 1997; 38(6):1074–1081.
die negativen Auswirkungen dieser Mutation auf die OPSIN-Proteinfaltung entdeckt:
Winderickx J, Sanocki E, Lindsey DT, Teller DY, Motulsky AG, Deeb SS. Defective colour vision associated with a missense mutation in the human green visual pigment gene. Nature Genetics. 1992;1:251–256. PMID: 1302020 .
und ihre Inzidenz festgestellt wurde, die bei hellhäutigen Personen bei ca. 2% liegt.
Informationen über seltene Mutation des Typs LIAVA finden sich hier:
Carroll J1, Neitz M, Hofer H, Neitz J, Williams DR., ‘Functional photoreceptor loss revealed with adaptive optics: an alternate cause of color blindness.’ Proc Natl Acad Sci U S A. 2004 Jun. PMID: 15148406.
Mizrahi-Meissonnier L., Merin S., Banin E., Sharon D., ‚Variable retinal phenotypes caused by mutations in the X-linked photopigment gene array‘. Invest. Ophthalmol. Vis. Sci. 2010 Aug;51(8):3884-92. PMID: 20220053.
Neitz M, Carroll J, Renner A, et al. ‚Variety of genotypes in males diagnosed as dichromatic on a conventional clinical anomaloscope‘. Vis Neurosci. 2004;21:205–216. PMID: 15518190.
Crognale MA, Fry M, Highsmith J, et al. ‚Characterization of a novel form of X-linked incomplete achromatopsia‘. Vis Neurosci. 2004; 21:197–203. PMID: 15518189.
Zu den ersten, weiter zurückliegenden Forschungsarbeiten über Blauzapfenmonochromasie zählen:
Huddart, J. ‚An account of persons who could not distinguish colours‘. Phil. Trans. Roy. Soc. 67: 260 only, 1777.
Sloan, L. L. ‚Congenital achromatopsia: a report of 19 cases‘. J. Ophthal. Soc. Am. 44: 117-128, 1954. PMID: 13131176.
Alpern, M., Falls, H. F., Lee, G. B. ‚The enigma of typical total monochromacy‘. Am. J. Ophthal. 50: 996-1012, 1960. PMID: 13682677.
Blackwell, H. R., Blackwell, O. M. Rod and cone receptor mechanisms in typical and atypical congenital achromatopsia. Vision Res. 1: 62-107, 1961. https://doi.org/10.1016/0042-6989(61)90022-0.
Fleischman, J. A., O’Donnell, F. E. Jr. ‘Congenital X-linked incomplete achromatopsia. Evidence for slow progression, carrier fundus findings, and possible genetic linkage with glucose-6-phosphate dehydrogenase locus’. Arch Ophthalmol 1981;99:468-472. PMID: 6971088.
Lewis, R. A., Holcomb, J. D., Bromley, W. C., Wilson, M. C., Roderick, T. H., Hejtmancik, J. F. ‘Mapping X-linked ophthalmic diseases: III. Provisional assignment of the locus for blue cone monochromacy to Xq28′. Arch. Ophthal. 105: 1055-1059, 1987. PMID: 2888453.
Erforschung der Zapfendystrophie, einer degenerativen Erkrankung durch Punktmutation in beiden Genen OPN1LW und OPN1MW:
Gardner JC, Webb TR, Kanuga N, Robson AG, Holder GE, Stockman A, Ripamonti C, Ebenezer ND, Ogun O, Devery S, Wright GA, Maher ER, Cheetham ME, Moore AT, Michaelides M and Hardcastle AJ,’X-Linked Cone Dystrophy Caused by Mutation of the Red and Green Cone Opsins‘.The American Journal of Human Genetics 87, 26–39, July 9, 2010. PMID: 20579627.
Schließlich seien auch einige Arbeiten genannt, die eine Übersicht über das Thema geben:
Neitz J., Neitz M. ‘The genetics of normal and defective color vision’. 2011 Review. Vision Research. PMID: 21167193.
Deeb, S.S. ‘Molecular Genetics of colour vision deficiencies’. Clinical and Experimental Optometry 87.4 – 5 July 2004. PMID:15312026.